[Messageboard]
![]()
|
|
Changes in the PfEMP protein on the malarial cell membrane :
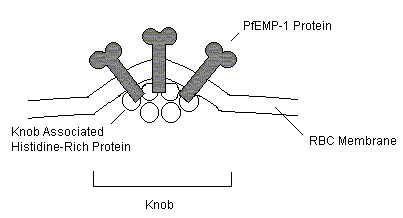
The ability of Plasmodium falciparum to reinfect previously exposed individuals is due to the existence of phenotypically variant strains of the parasite in endemic areas. The epidemiology of malaria is heavily dependent upon this variance. Clinical studies have shown thatimmunity to malaria requires repeated infections and is slow to develop, and that children under ten years of age are the most susceptible to grave illness and fatality. This pathology is explained by the diversity of P. Falciparum strains and the variant allelic forms of parasite proteins that they produce. Cerebral malaria, a fatal form of the disease, is caused by the aggregation of infected red blood cells adhering to and blocking capillaries in the brain. Adults who have been exposed to variant strains of the P. Falciparum and are able produce a sufficient humoral response to parasite proteins involved in parasite cytoadherence. Children, however, have not been immunologically exposed to this diverse body of parasite proteins. Their inability to mount a strong immune response to variant strains can, in extreme situations, result in fatality. Antigenic diversity of parasite strains is a product of genetic recombination during both the sexual, vector phase and the asexual amplification phase during human red blood cell infection. Individual Parasite Diversity Antigenic diversity at the level of the individual parasite occurs during the course of infection of a given individual. Variation occurs in the surface antigens of infected red blood cells during the erythocytic schizogany phase of the parasite life cycle. The sequestration of the malaria parasites in human red blood cells during this phase of the cycle poses an extremely complex method for evasion to the host immunity. First, infected RBCs do not induce CTL response due to their lackof MHC expression. Second, parasite derived particles exposed on the surface of the cell are highly variable, leading to the inability of the immune response to produce adequate memory to the antigens. These molecules are associated with PfEMP-1 proteins and undergo high clonal variation (2% per generation, in culture).
|