[Messageboard]
![]()
|
|
The complete and detailed Malaria vaccine review: Note: This section was written by Dr. Stephanie James and Dr.Louis Miller for NIAID (a US govt. agency) and I totally disown any adverse effects resulting from reading this article.. (also read "Hyper-electrosis Cerebri" , posted on this fortnight's opening page... ) The
brutal fact is that this article is too difficult to be completely understood by
medical graduates. If someone was to talk to me about Gp-120, I might
think for a while and might even realise that the particular person was talking
about HIV / AIDS. However, PfEMP-1 makes about as much sense to me as my
mother.. :) just kidding... You
might want to read the table below and skip the rest of the section. If however,
you are fascinated by the malarial parasite and are seriously considering
marrying the damn bug, you might want to read the rest of it too. Malaria
: Opportunities and Obstacles (Published by the National Academy press)
The
Understandable Table
Introduction:
Several
early vaccine candidates, many based on the circumsporozoite (CS) protein, the
dominant surface antigen of the sporozoite stage, progressed into Phase I/II
clinical trials but were halted by problems of low immunogenicity and efficacy
or, in some cases, by reactogenicity. Overall, those candidates that have
proceeded to trials have generally had some form of corporate co-sponsorship.
Only one candidate vaccine, SPf66, based on antigens from both merozoite and
sporozoite stages, has undergone extensive field trials. Efficacy was reported
in several early clinical trials in South America, and one in Africa, but
results from subsequent trials in Africa and Southeast Asia were not as
promising. These
results underscore the need not only to identify the right antigenic components
for a vaccine, but also to find presentation and delivery methods that induce
qualitatively and quantitatively appropriate immune responses.
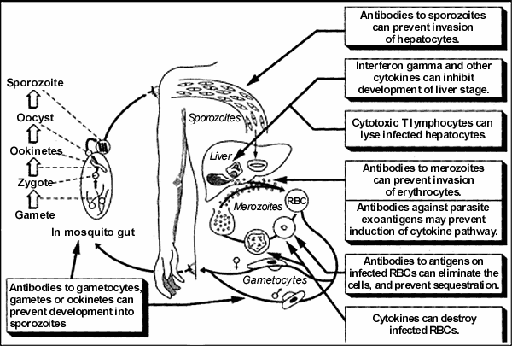
Immunology : Experimental observations indicate that protective immunity may involve multiple different immune responses, both humoral and cellular (Fig. 1). The focus has been largely on induction of antibodies in the case of blood-stage and transmission-blocking vaccines. Pre-erythrocytic
vaccine developers initially focused on induction of antibodies, then on CD8+
cytotoxic T lymphocytes (CTL), and now additionally on T cell derived cytokines.
Interferon gamma (IFNg) and other cytokines appear to play a role in the
elimination of liver stage parasites, possibly through induction of mediators
such as nitric oxide that kill parasites within hepatocytes (Fig. 1). Figure
1 — Possible Mechanisms of Host Defense Against Malaria
To
date, no pattern of immune response fully predictive of protection has been
identified or validated. Naturally occurring immunity wanes rapidly in the
absence of ongoing parasite exposure, and protection has been similarly
short-lived in those few subunit vaccine trials that have demonstrated
measurable efficacy. Such a vaccine might be useful for travelers. Unless new
technologies can be found to improve the longevity of vaccine-induced
resistance, however, it is likely that a vaccine to be used in endemic areas
will need to take advantage of natural boosting provided by ongoing parasite
exposure in order to provide long-lived protection. In
natural infection, unregulated immune responses may contribute to the
pathogenesis of disease. For example, an association of cerebral malaria with
high plasma levels of pro-inflammatory cytokines has been reported. Thus, the
potential for enhanced immunopathogenesis must also be taken into account in
vaccine development efforts.
St To
date, most of the effort on vaccine development has focused on P. falciparum for
several reasons: While
P. vivax has a wider geographic distribution than P. falciparum, including in
emerging economies such as Southeast Asia, India and Brazil, work on vaccine
development has been impeded by several technical obstacles, such as the
difficulty of culturing the parasite in vitro. Because
it is impossible to be all-encompassing in the review of current research on
malaria vaccines within the limitations of this document, the summary below aims
only to illustrate some of the newest approaches to vaccine development.
1)Recombinant
vaccines A.
The Walter Reed Army institute vaccine: This collaboration has led to development of the CS-based RTS,S vaccine, which in combination with SBAS2 adjuvant repeatedly induced protection of volunteers in a Phase IIa trial. Subsequent re-challenge of volunteers revealed that protection waned substantially by 6 months after the last immunization. The first field trial of RTS,S/SBAS2, conducted by the Medical Research Council in The Gambia, reported 65% efficacy in adult males in a regions of intense transmission where both homologous and heterologous P. falciparum strains are prevalent. Efficacy persisted for 2 months and diminished afterward. In
partnership with SBBio, USAID, the Naval Medical Research Center (NMRC) and
others, WRAIR is conducting preclinical, clinical, and field trials to: . B)The
NIAID Malaria Vaccine Development Unit (MVDU) The
MVDU has facilities for protein expression in a variety of recombinant systems
as well as subsequent process development. Once produced and purified,
blood-stage antigens are being tested in Aotus monkeys to identify the most
promising candidates. Transmission-blocking
vaccines under development at the MVDU include Pfs25 and Pvs25, sexual stage (ookinete)
antigens expressed by P. falciparum and P. vivax, respectively. Clinical grade
Pfs25 has been produced and a plan is underway for evaluating these antigens for
safety and immunogenicity in clinical trials later this year. Clinical grade
Pvs25 is also being prepared for Phase I testing. C.
New York University: D.
The CDC malaria vaccine program is focused on development of
multivalent, multistage vaccine formulations that contain a series of antigenic
domains from all of the developmental stages of the parasite. The multivalent,
multistage malaria vaccine development strategy, which is aimed at inducing
"multiple layers" of long-lasting, effective immunity, takes into
consideration the immunogenicity and genetic diversity of antigenic fragments
contained in stage-specific proteins.
2)
DNA
vaccine and prime-boost approaches The Naval Medical Research Center (NMRC), in collaboration with Vical, Inc., USAID, Aventis Pasteur, Entremed, Inc., and multiple investigators around the world, has initiated a DNA-based malaria vaccine development effort called the "Multi-Stage DNA Vaccine Operation" or "MuStDO" Researchers
at Oxford are assessing the safety, immunogenicity and efficacy
of a DNA prime-MVA (modified vaccinia virus Ankara) boost regime in healthy
volunteers. The insert, which is the same in the DNA and the MVA components, is
a polyepitope string fused to the entire TRAP antigen (Thrombospondin-Related
Anonymous Protein, also known as Sporozoite Surface Protein 2, SSP2, which is
expressed primarily by sporozoites and liver stage parasites). The delivery of
DNA by intramuscular route and by a needleless delivery device into the skin is
being compared. MVA is delivered intradermally. Initial studies of both DNA and
MVA vaccines established adequate safety and immunogenicity. The first
prime-boost studies of volunteers were initiated in December, 1999, and are
ongoing, with challenge studies anticipated later this year. Plans are underway
to test both DNA and MVA vaccines in The Gambia in late 2000.
3)
Transgenic vaccines Genzyme
Transgenics Corp. and NIAID have established a collaboration for preclinical
development of an MSP-1-based vaccine in transgenic animals. The
42 kd fragment of MSP1 has been produced in milk of transgenic mice, and a
purification process is now under development. If the purified product can be
shown to protect Aotus monkeys, it will provide supporting data supporting the
development of other transgenic animals such as goats.
4)
Genomic and proteomic approaches : In
1996, a collaborative international effort was undertaken to sequence the
complete genome of P. falciparum. To
date, two of the parasite's 14 chromosomes have been completely sequenced and
the sequences of many of the remaining chromosomes are nearing completion.
Although there have been significant technical hurdles in sequencing this A-T
rich organism, it is now estimated that essentially all of the parasites
approximately 6000 genes are available in existing databases. Thus, the malaria
community and vaccine developers have access to virtually all of the genes
encoding the antigens and proteins expressed by this organism. Although not
validated, computer algorithms are being conceived to identify genes whose
expressed products are potential candidate vaccine antigens by virtue of their
predicted cell surface localization, stage specific expression, or structural
features that may interact with components of the human host immune system to
initiate a protective response. Accompanying the development of such
computational methods are high throughput methods (DNA chips and microarrays,
and proteomics) of differential gene and protein expression to identify and
characterize antigens that may be appropriately exposed to the immune system. A
challenge for malaria investigators is the limiting amounts of material that are
available for certain stages of the parasite's life cycle, especially the liver
stages. Investigators
are developing strategies to identify genes whose products are essential to
parasite survival and/or contribute to disease manifestations. Interfering
with the function of these gene products with targeted immune responses is
another approach to malaria vaccine development. New tools of molecular
genetics, e.g. efficient and global gene knockouts, are needed to make this
approach truly feasible and economically viable.
| |||||||||||||||||||||||||||||||||